

Italiano

English
|
| Italiano | English |

| Alternative all'invecchiamento
[Traduzione della pagina:
"http://www.programmed-aging.org/theory-2/alternatives_to_aging.htm" dal sito: "http://www.programmed-aging.org"] |
Esistono tre principali forme di curva di sopravvivenza alternative all'invecchiamento (progressivo declino della fitness
in correlazione con l'etŕ) (Finch 1990), che non sono descrivibili con la formula di Weibull:
Prima alternativa: Morte chiaramente programmata
Tipo 1: Rapido invecchiamento e morte dopo la riproduzione
Esempi: Molte specie di salmoni (Finch 1990, pp. 83-87), aracnidi [ibidem,
p. 76], molluschi [p. 78], lamprede [p. 80], alcune specie di piccoli marsupiali [p. 95] e roditori [p. 97],
molte piante [p. 98], etc.
Tipo 2: Morte obbligatoria per mancanza di parti anatomiche nello stadio adulto
“L'afagia in conseguenza della mancanza di parti della bocca o degli organi digestivi č molto comune negli stadi adulti degli
insetti … ed č il fattore limitante della durata della vita della fase adulta in molte specie con breve longevitŕ” [ibidem, p. 49], etc.
Tipo 3: Morte obbligatoria della madre in relazione alla nascita della prole
E.g.: “il nematode Ascaris nigrovenosa … il giovane uccide la madre fuoriuscendo attraverso la parete
del suo corpo” [ibidem, p. 102], etc.
Seconda alternativa: Senescenza non rilevabile (longevitŕ indeterminata), senescenza negativa
Tipo 1: Senescenza non rilevabile
Si definisce che una specie ha una senescenza non rilevabile quando:
“(1) il tasso di mortalitŕ dopo la maturitŕ rimane relativamente costante in relazione all'etŕ; o (2) nella fase matura
non vi sono rilevabili compromissioni funzionali in relazione all'etŕ.” (Finch 1990, p. 208)
Esempi: “Certe conifere (gimnosperme) presentano enormi longevitŕ di molte migliaia di anni allo
stato naturale” [ibidem, p. 209]; “numerosi artropodi mostrano crescita indeterminata e nessun segno di
invecchiamento” [p. 213], molte specie di rockfish [http://www.agelessanimals.org/index.htm], “[Vi sono]
spugne e altri organismi che si riproducono in modo asessuale in cui molte o tutte le cellule continuano a proliferare
indefinitamente e danno la capacitŕ per una considerevole rigenerazione di organi danneggiati; tali specie tendono
ad avere longevitŕ indefinite” [p. 61], etc.
Tipo 2: Senescenza negativa
In alcuni casi il progressivo crescere della massa corporea, che riduce la possibilitŕ di essere predati,
o altri fattori, riducono il tasso di mortalitŕ che altrimenti sarebbe presumibilmente
stabile (Comfort 1979) (Congdon et
al. 2003) (Vaupel et al. 2004).
“Senescenza negativa”, un termine potenzialmente fuorviante, dovrebbe essere considerato un caso particolare
di senescenza non rilevabile.
Terza alternativa: Mortalitŕ costante allo stato selvatico e mortalitŕ crescente a partire da etŕ
non esistenti allo stato naturale
La mancanza di turnover cellulare significa che cellule ed organi non possono mantenersi per sempre a causa
dell'azione di fattori meccanici e biochimici che progressivamente li danneggiano. E.g.: “gli insetti [sono]
a rischio per danni che indeboliscono cellule ed organi insostituibili, in particolare le loro fragili ali
e l'esoscheletro, cosa che Comfort (1979) ha chiamato invecchiamento mecchanico” (Finch 1990).
Se questa mancanza di turnover cellulare causa danno organico / cellulare al di sopra di un livello critico
a partire da etŕ non esistenti in condizioni naturali, la mancanza di turnover cellulare č irrilevante
per la selezione naturale.
La mosca Drosophila melanogaster ed il nematode Caenorhabditis elegans, entrambi composti di cellule
senza turnover (Finch 1990) (Arking 1998) sono esempi famosi di questo caso.
Queste due specie mostrano in laboratorio una mortalitŕ crescente in correlazione con l'etŕ illustrata
nella fig. 1 (ridisegnata da (Finch 1990)) e nella fig. 2 (ridisegnata da
(Finch & Hayflick 1977).
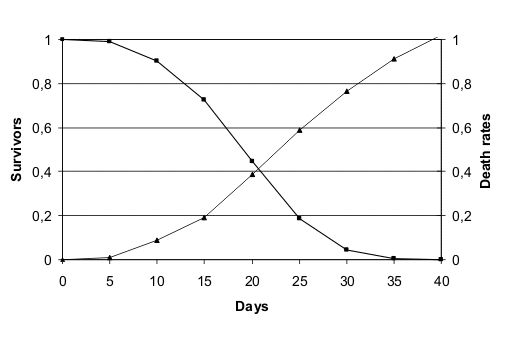 Curva di sopravvivenza in laboratorio di Caenorhabditis elegans. | 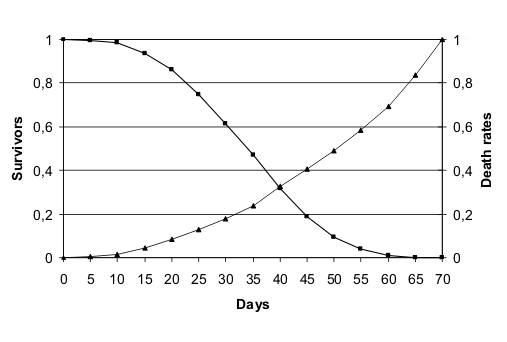 Curva di sopravvivenza in laboratorio di Drosophila melanogaster. |
www.programmed-aging.org
sponsored by AZINET LLC © 2009